
The Golgi apparatus, also known as Golgi complex, it is a membranous cell organelle formed by a set of flat vesicles stacked together; these sacks have fluid inside. Found in a wide variety of eukaryotes, including animals, plants, and fungi.
This organelle is responsible for the processing, packaging, classification, distribution and modification of proteins. In addition, it also has a role in the synthesis of lipids and carbohydrates. On the other hand, in plants the synthesis of the components of the cell wall occurs in the Golgi apparatus..
The Golgi apparatus was discovered in 1888, while nerve cells were being studied; its discoverer, Camillo Golgi, won the Nobel Prize. The structure is detectable by silver chromate staining.
At first the existence of the organ was doubtful for the scientists of the time and they attributed the Golgi observations to simple artifacts, products of the techniques used..
Article index
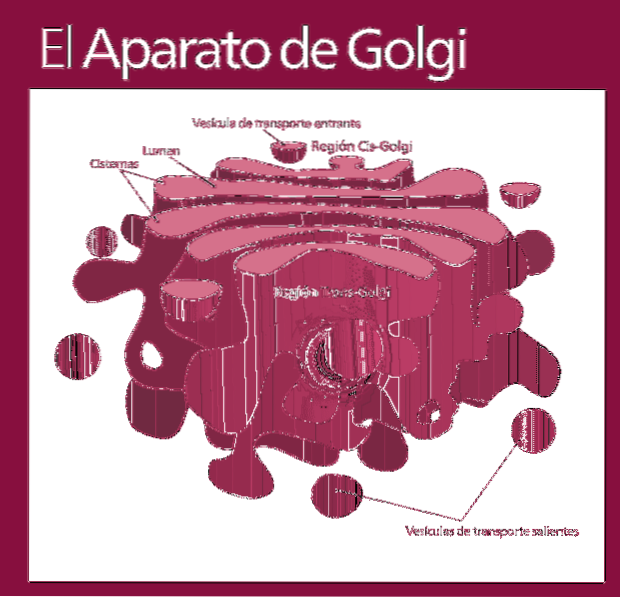
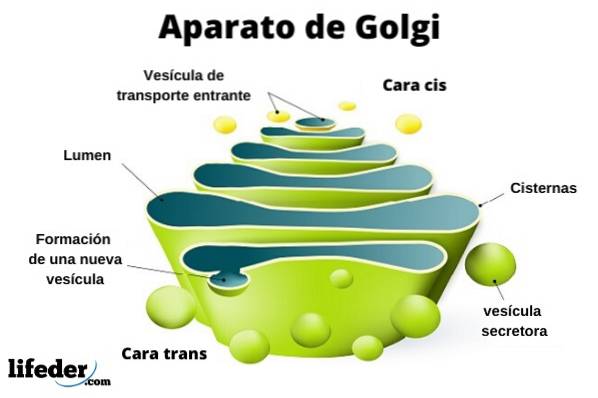
The Golgi apparatus is a eukaryotic organelle of a membranous nature. It resembles bags in piles, although the organization may vary depending on the cell type and the organism. It is responsible for the modification of proteins after translation.
For example, some carbohydrate can be added to form a glycoprotein. This product is packaged and distributed to the cell compartment where it is required, such as the membrane, lysosomes or vacuoles; it can also be sent outside the cell. It also participates in the synthesis of biomolecules.
The cytoskeleton (specifically actin) determines its location, and generally the complex is located in an area of the interior of the cell close to the nucleus and the centrosome.
The Golgi complex is made up of a set of flat, fenestrated, disc-shaped sacs, called Golgian cisterns, of varying thickness..
These sacks are stacked, in groups of four or six cisterns. In a mammalian cell you can find between 40 and 100 batteries connected to each other.
The Golgi complex presents an interesting characteristic: there is polarity in terms of structure and also of function.
You can distinguish between the cis face and the trans face. The first is related to the entry of proteins and is located near the endoplasmic reticulum. The second is the exit face or product secretion; they are formed by one or two tanks that have a tubular shape.
Along with this structure are vesicles that make up the transport system. The stacks of sacks are joined together in a structure reminiscent of the shape of a bow or a date.
In mammals, the Golgi complex is fragmented into several vesicles during cell division processes. The vesicles pass to the daughter cells and again take the traditional form of the complex.
The organization of the complex is not common in all groups of organisms. In some cell types the complex is not structured as sets of cisterns stacked in groups; on the contrary, they are located individually. An example of this organization is that of the fungus Saccharomyces cerevisiae.
In some single-celled organisms, such as toxoplasma or trypanosoma, the presence of only one membranous pile has been reported.
All these exceptions indicate that the stacking of the structures is not essential to fulfill their function, although the proximity between the bags makes the transport process much more efficient..
Similarly, some basal eukaryotes lack these cisternae; for example, mushrooms. This evidence supports the theory that the apparatus appeared in a later lineage than the first eukaryotes..
Functionally, the Golgi complex is divided into the following compartments: the cis network, the stacked sacs -which in turn are divided into the middle and trans subcompartment- and the trans network.
The molecules to be modified enter the Golgi complex following the same order (cis network, followed by subcompartments to be finally excreted in the trans network).
Most reactions occur in the most active zones: the trans and middle subcompartments.
The Golgi complex has as its main function the post-translational modification of proteins thanks to the enzymes they have inside.
These modifications include processes of glycosylation (addition of carbohydrates), phosphorylation (addition of a phosphate group), sulfation (addition of a phosphate group) and proteolysis (degradation of proteins).
Furthermore, the Golgi complex is involved in the synthesis of specific biomolecules. Each of its functions is described in detail below:
In the Golgi apparatus, the modification of a protein to a glycoprotein occurs. The typical acidic pH of the interior of the organelle is critical for this process to occur normally..
There is a constant exchange of materials between the Golgi apparatus with the endoplasmic reticulum and the lysosomes. In the endoplasmic reticulum, proteins also undergo modifications; these include the addition of an oligosaccharide.
When these molecules (N-oligosaccharides) enter the Golgi complex, they receive a series of additional modifications. If the destiny of this molecule is to be carried outside the cell or to be received in the plasma membrane, special modifications occur.
These modifications include the following steps: the removal of three mannose residues, the addition of N-acetylglucosamine, the removal of two mannoses, and the addition of fucose, two additional N-acetylglucosamine, three galactose, and three sialic acid residues..
In contrast, the proteins that are destined for lysosomes are modified in the following way: there is no removal of mannoses as an initial step; instead, phosphorylation of these residues occurs. This step occurs in the cis region of the complex.
Next, the N-acetylglucosamine groups are removed leaving the mannoses with the phosphate added to the oligosaccharide. These phosphates indicate that the protein must be specifically targeted to lysosomes..
The receptors that are responsible for recognizing the phosphates that indicate their intracellular fate are located in the trans network.
In the Golgi complex, the synthesis of glycolipids and sphingomyelin occurs, using ceramide (previously synthesized in the endoplasmic reticulum) as the molecule of origin. This process is contrary to that of the rest of the phospholipids that make up the plasma membrane, which are derived from glycerol..
Sphingomyelin is a class of sphingolipid. It is an abundant component of mammalian membranes, particularly nerve cells, where they are part of the myelin sheath.
After their synthesis, they are transported to their final location: the plasma membrane. Their polar heads are located towards the outside of the cell surface; these elements have a specific role in cell recognition processes.
In plant cells, the Golgi apparatus contributes to the synthesis of the polysaccharides that make up the cell wall, specifically hemicellulose and pectins. Through vesicular transport, these polymers are carried outside the cell.
In vegetables, this step is crucial and approximately 80% of the activity of the reticulum is assigned to the synthesis of polysaccharides. In fact, hundreds of these organelles have been reported in plant cells..
The different biomolecules - proteins, carbohydrates and lipids - are transferred to their cellular destinations by the Golgi complex. Proteins have a kind of "code" that is responsible for informing the destination to which it belongs.
They are transported in vesicles that leave the trans network and travel to the specific cell compartment.
Proteins can be carried to the membrane by a specific constitutive pathway. That is why there is a continuous incorporation of proteins and lipids to the plasma membrane. Proteins whose final destination is the Golgi complex are retained by this.
In addition to the constitutive pathway, other proteins are destined for the exterior of the cell and occur through signals from the environment, be it hormones, enzymes or neurotransmitters..
For example, in pancreatic cells, digestive enzymes are packaged in vesicles that are only secreted when food is detected..
Recent research reports the existence of alternative pathways for membrane proteins that do not pass through the Golgi apparatus. However, these pathways bypass "Unconventional" are debated in the literature.
There are five models to explain protein trafficking in the apparatus. The first involves the traffic of material between stable compartments, each one has the necessary enzymes to fulfill specific functions. The second model involves the maturation of the cisterns progressively.
The third also proposes the maturation of the bags but with the incorporation of a new component: the tubular transport. According to the model, tubules are important in traffic in both directions.
The fourth model proposes that the complex works as a unit. The fifth and last model is the most recent and maintains that the complex is divided into different compartments.
In certain cell types the Golgi complex has specific functions. The cells of the pancreas have specialized structures for the secretion of insulin.
The different blood types in humans are an example of differential glycosylation patterns. This phenomenon is explained by the presence of different alleles that code for glucotransferase.



Yet No Comments