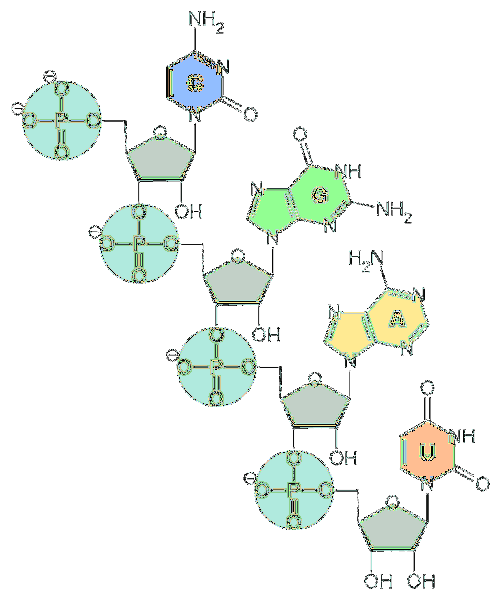
The RNA or RNA (ribonucleic acid) is a type of nucleic acid present in eukaryotic and prokaryotic organisms and in viruses. It is a nucleotide polymer that contains four types of nitrogenous bases in its structure: adenine, guanine, cytosine and uracil.
RNA is generally found as a single band (except in some viruses), in a linear fashion or in a series of complex structures. In fact, RNA has a structural dynamism that is not observed in the DNA double helix. Different types of RNA have very varied functions.
Ribosomal RNAs are part of ribosomes, the structures in charge of protein synthesis in cells. Messenger RNAs function as intermediaries and carry genetic information to the ribosome, which translates the message from a nucleotide sequence to an amino acid sequence.
Transfer RNAs are responsible for activating and transferring the different types of amino acids -20 in total- to the ribosomes. There is a transfer RNA molecule for each amino acid that recognizes the sequence in messenger RNA.
In addition, there are other types of RNA that are not directly involved in protein synthesis and participate in gene regulation..
Article index
The fundamental units of RNA are nucleotides. Each nucleotide is made up of a nitrogenous base (adenine, guanine, cytosine, and uracil), a pentose, and a phosphate group..
Nitrogen bases are derived from two fundamental compounds: pyrimidines and purines..
The bases derived from purines are adenine and guanine and the bases derived from pyrimidines are cytosine and uracil. Although these are the most common bases, nucleic acids can also have other types of bases that are less common..
As for pentose, they are units of d-ribose. For this reason, the nucleotides that make up RNA are called "ribonucleotides".
Nucleotides are linked together by chemical bonds involving the phosphate group. To form them, the phosphate group at the 5 'end of a nucleotide is attached to the hydroxyl group (-OH) at the 3' end of the next nucleotide, thus creating a phosphodiester-like bond..
Along the nucleic acid chain, the phosphodiester bonds have the same orientation. Therefore, there is a polarity of the strand, distinguishing between the 3 'and the 5' end.
By convention, the structure of nucleic acids is represented with the 5 'end on the left and the 3' end on the right.
The RNA product of DNA transcription is a single stranded band that turns to the right, in a helical conformation by the stacking of bases. The interaction between purines is much greater than the interaction between two pyrimidines, due to their size.
In RNA, it is not possible to speak of a traditional and reference secondary structure, such as the double helix of DNA. The three-dimensional structure of each RNA molecule is unique and complex, comparable to that of proteins (logically, we cannot globalize the structure of proteins).
There are weak interactions that contribute to the stabilization of RNA, particularly base stacking, where the rings are located one on top of the other. This phenomenon also contributes to the stability of the DNA helix..
If the RNA molecule finds a complementary sequence, they will be able to couple and form a double-stranded structure that turns to the right. The predominant form is type A; As for the Z forms, they have only been evidenced in the laboratory, while the B form has not been observed.
Generally, there are short sequences (such as UUGG) that are located at the end of the RNA and have the particularity of forming loops stable. This sequence participates in the folding of the three-dimensional structure of RNA.
Furthermore, hydrogen bonds can form at other sites than the typical base pairings (AU and CG). One of these interactions occurs between the 2'-OH of ribose with other groups.
Elucidating the various structures found in RNA has served to demonstrate the multiple functions of this nucleic acid..
There are two classes of RNA: informational and functional. The first group includes RNAs that participate in protein synthesis and function as intermediaries in the process; informational RNAs are messenger RNAs.
In contrast, RNAs belonging to the second class, the functional ones, do not give rise to a new protein molecule and the RNA itself is the final product. These are transfer RNAs and ribosomal RNAs..
In mammalian cells, 80% of RNA is ribosomal RNA, 15% is transfer RNA, and only a small portion corresponds to messenger RNA. These three types work cooperatively to achieve protein biosynthesis..
There are also small nuclear RNAs, small cytoplasmic RNAs, and microRNAs, among others. Each of the most important types will be described in detail below:
In eukaryotes, DNA is confined to the nucleus, while protein synthesis occurs in the cytoplasm of the cell, where ribosomes are found. Due to this spatial separation there must be a mediator that carries the message from the nucleus to the cytoplasm and that molecule is the messenger RNA..
Messenger RNA, abbreviated mRNA, is an intermediate molecule that contains the information encoded in DNA and that specifies an amino acid sequence that will give rise to a functional protein..
The term messenger RNA was proposed in 1961 by François Jacob and Jacques Monod to describe the portion of RNA that transmitted the message from DNA to ribosomes..
The process of synthesis of an mRNA from the DNA strand is known as transcription and it occurs differentially between prokaryotes and eukaryotes..
Gene expression is governed by several factors and depends on the needs of each cell. Transcription is divided into three stages: initiation, elongation, and termination..
The DNA replication process, which occurs in each cell division, copies the entire chromosome. However, the transcription process is much more selective, it only deals with processing specific segments of the DNA strand and does not require a primer..
On Escherichia coli -the best studied bacterium in life sciences - transcription begins with the unwinding of the DNA double helix and the transcription loop is formed. The enzyme RNA polymerase is responsible for synthesizing RNA and, as transcription continues, the DNA strand returns to its original shape.
Transcription is not initiated at random sites on the DNA molecule; there are specialized sites for this phenomenon, called promoters. On E. coli RNA polymerase is coupled a few base pairs above the target region.
The sequences where transcription factors are coupled are quite conserved between different species. One of the best known promoter sequences is the TATA box.
On elongation, the RNA polymerase enzyme adds new nucleotides to the 3'-OH end, following the 5 'to 3' direction. The hydroxyl group acts as a nucleophile, attacking the alpha phosphate of the nucleotide to be added. This reaction releases a pyrophosphate.
Only one of the DNA strands is used to synthesize the messenger RNA, which is copied in the 3 'to 5' direction (the antiparallel form of the new RNA strand). The nucleotide to be added must comply with base pairing: U pairs with A, and G with C.
RNA polymerase stops the process when it finds regions rich in cytosine and guanine. Eventually, the new messenger RNA molecule is separated from the complex.
In prokaryotes, a messenger RNA molecule can code for more than one protein.
When an mRNA exclusively codes for a protein or polypeptide, it is called monocistronic mRNA, but if it codes for more than one protein product, the mRNA is polycistronic (note that in this context the term cistron refers to the gene).
In eukaryotic organisms, the vast majority of mRNAs are monocistronic and the transcriptional machinery is much more complex in this lineage of organisms. They are characterized by having three RNA polymerases, denoted I, II and III, each with specific functions.
The I is in charge of synthesizing the pre-rRNAs, the II synthesizes the messenger RNAs and some special RNAs. Finally, III is responsible for transfer RNAs, ribosomal 5S, and other small RNAs..
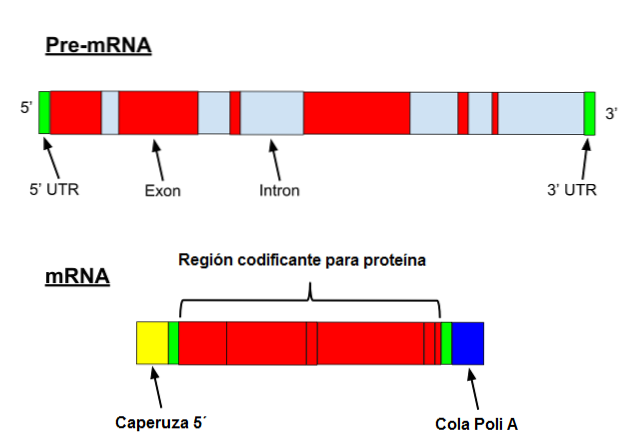
Messenger RNA undergoes a series of specific modifications in eukaryotes. The first involves adding a "cap" to the 5 'end. Chemically, the cap is a 7-methylguanosine residue attached to the end by a 5 ', 5'-triphosphate bond..
The function of this zone is to protect RNA from possible degradation by ribonucleases (enzymes that break down RNA into smaller components).
In addition, removal of the 3 'end occurs and 80 to 250 adenine residues are added. This structure is known as the polyA "tail" and serves as a binding site for various proteins. When a prokaryote acquires a polyA tail, it tends to stimulate its degradation..
On the other hand, this messenger is transcribed with introns. Introns are DNA sequences that are not part of the gene but that "interrupt" that sequence. Introns are not translated and therefore must be removed from the messenger.
Most vertebrate genes have introns, with the exception of genes that code for histones. Similarly, the number of introns in a gene can vary from a few to tens of these..
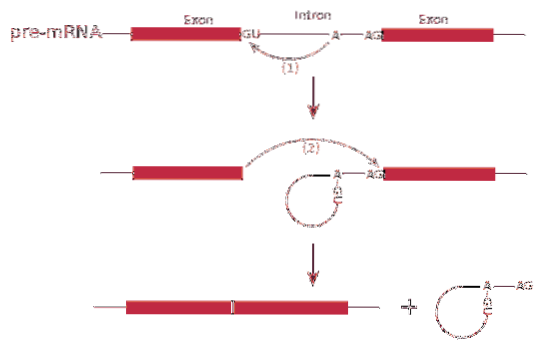
The splicing RNA or splicing process involves the removal of introns in messenger RNA.
Some introns found in nuclear or mitochondrial genes can carry out the process of splicing without the help of enzymes or ATP. Instead, the process is carried out by transesterification reactions. This mechanism was discovered in the ciliated protozoan Tetrahymena thermophila.
In contrast, there is another group of messengers who are not capable of mediating their own splicing, so they need additional machinery. A fairly large number of nuclear genes belongs to this group..
The process of splicing it is mediated by a protein complex called a spliceosome or splicing complex. The system is made up of specialized RNA complexes called small nuclear ribonucleoproteins (RNP)..
There are five types of RNP: U1, U2, U4, U5 and U6, which are found in the nucleus and mediate the process of splicing.
The splicing can produce more than one type of protein - this is known as splicing alternative-, since the exons are arranged differentially, creating varieties of messenger RNAs.
Ribosomal RNA, abbreviated rRNA, is found in ribosomes and participates in protein biosynthesis. Therefore, it is an essential component of all cells.
Ribosomal RNA associates with protein molecules (approximately 100) to give rise to ribosomal presubunits. They are classified depending on their sedimentation coefficient, denoted by the letter S for Svedberg units..
A ribosome is made up of two parts: the major subunit and the minor subunit. Both subunits differ between prokaryotes and eukaryotes in terms of the coefficient of sedimentation..
Prokaryotes have a large 50S subunit and a small 30S subunit, while in eukaryotes the large subunit is 60S and the small 40S.
The genes that code for ribosomal RNAs are in the nucleolus, a particular area of the nucleus that is not bounded by a membrane. Ribosomal RNAs are transcribed in this region by RNA polymerase I.
In cells that synthesize large amounts of proteins; the nucleolus is a prominent structure. However, when the cell in question does not require a high number of protein products, the nucleolus is an almost imperceptible structure.
The 60S large ribosomal subunit is associated with the 28S and 5.8S fragments. Regarding the small subunit (40S), it is associated with 18S.
In higher eukaryotes, the pre-rRNA is encoded in a 45S transcriptional unit, which involves RNA polymerase I. This transcript is processed into mature 28S, 18S and 5.8S ribosomal RNAs..
As synthesis continues, the pre-rRNA associates with different proteins and forms ribonucleoprotein particles. It undergoes a series of subsequent modifications that include methylation of the 2'-OH group of ribose and the conversion of uridine residues to pseudouridine..
The region where these changes will occur is controlled by more than 150 small nucleolar RNA molecules, which have the ability to couple to pre-rRNA..
Contrary to the rest of the pre-rRNAs, 5S is transcribed by RNA polymerase III in the nucleoplasm and not inside the nucleolus. After being synthesized, it is taken to the nucleolus to assemble with the 28S and 5.8S, thus forming the ribosomal units.
At the end of the assembly process, the subunits are transferred to the cytoplasm through the nuclear pores.
It can happen that a messenger RNA molecule gives rise to several proteins at the same time, binding to more than one ribosome. As the translation process progresses, the end of the messenger becomes free and can be taken up by another ribosome, starting a new synthesis..
For this reason, it is common to find ribosomes grouped (between 3 and 10) in a single messenger RNA molecule, and this group is called polyribosome.
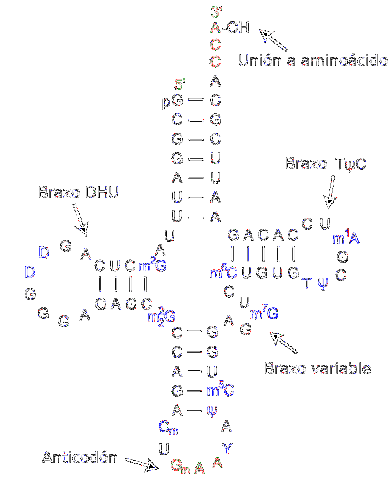
Transfer RNA is responsible for transferring amino acids as the protein synthesis process progresses. They are made up of approximately 80 nucleotides (compared to messenger RNA, it is a "small" molecule).
The structure has folds and crosses reminiscent of a three-armed shamrock. An adenylic ring is located at one of the ends, where the hydroxyl group of ribose mediates the union with the amino acid to be transported.
The different transfer RNAs combine exclusively with one of the twenty amino acids that make up proteins; in other words, it is the vehicle that carries the building blocks of proteins. The transfer RNA complex together with the amino acid is called aminoacyl-tRNA.
Furthermore, in the translation process - which occurs thanks to ribosomes - each transfer RNA recognizes a specific codon in the messenger RNA. When recognized, the corresponding amino acid is released and becomes part of the synthesized peptide.
To recognize the type of amino acid that must be delivered, the RNA has an "anticodon" located in the middle region of the molecule. This anticodon is capable of forming hydrogen bonds with the complementary bases present in messenger DNA..
MicroRNAs or miRNAs are a type of short, single-stranded RNA, between 21 and 23 nucleotides, whose function is to regulate the expression of genes. Since it is not translated into protein, it is often called non-coding RNA.
Like other types of RNA, microRNA processing is complex and involves a number of proteins.
MicroRNAs arise from longer precursors called mi-pri-RNAs, derived from the first transcript of the gene. In the nucleus of the cell, these precursors are modified in the microprocessor complex and the result is a pre-miRNA..
Pre-miRNAs are 70 nucleotide hairpins that continue to be processed in the cytoplasm by an enzyme called Dicer, which assembles the RNA-induced silencing complex (RISC) and finally the miRNA is synthesized..
These RNAs are capable of regulating the expression of genes, since they are complementary to specific messenger RNAs. By mating with their target, the miRNAs are able to suppress the messenger, or even degrade it. Consequently, the ribosome cannot translate said transcript..
One particular type of microRNA is small interfering RNAs (siRNAs), also called silencing RNAs. They are short RNAs, between 20 to 25 nucleotides, which hinder the expression of certain genes.
They are very promising instruments for research, since they make it possible to silence a gene of interest and thus study its possible function..
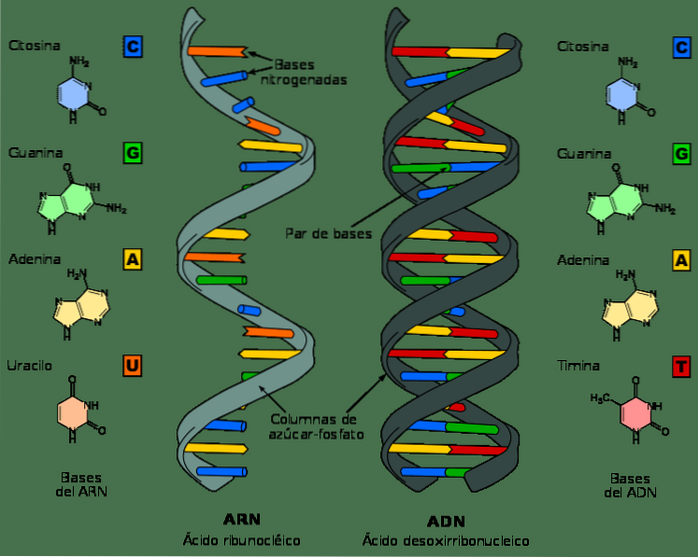
Although DNA and RNA are nucleic acids and can look very similar at first glance, they differ in several of their chemical and structural properties. DNA is a double-band molecule, while RNA is single-band.
As a result, RNA is a more versatile molecule and can take on a wide variety of three-dimensional shapes. However, certain viruses have double-band RNA in their genetic material..
In RNA nucleotides the sugar molecule is a ribose, while in DNA it is a deoxyribose, differing only in the presence of an oxygen atom.
The phosphodiester bond in the backbone of DNA and RNA is prone to undergo a slow hydrolysis process without the presence of enzymes. Under alkaline conditions, RNA hydrolyzes rapidly - thanks to the extra hydroxyl group - while DNA does not.
Similarly, the nitrogenous bases that make up nucleotides in DNA are guanine, adenine, thymine, and cytosine; in contrast, in RNA, thymine is replaced by uracil. Uracil can pair with adenine, in the same way as thymine in DNA.
RNA is the only known molecule capable of storing information and catalyzing chemical reactions at the same time; Therefore, several authors propose that the RNA molecule was crucial in the origin of life. Surprisingly, ribosome substrates are other RNA molecules.
The discovery of ribozymes led to the biochemical redefinition of "enzyme" -since previously the term was used exclusively for proteins with catalytic activity-, and helped support a scenario where the first forms of life used only RNA as genetic material.


Yet No Comments