
The hexokinase (HK) is the enzyme that catalyzes the first reaction of the glycolytic pathway in virtually all living organisms, both prokaryotes and eukaryotes. It is responsible for the transfer of a phosphoryl group to glucose, producing glucose-6P, although it can also phosphorylate other sugars hexoses (6 carbon atoms).
This enzyme is classified within one of the two families of glucose kinase enzymes (enzymes that transfer phosphoryl groups to substrates such as glucose): the hexokinase (HK) family, whose members are subdivided into three different groups known as the HK group, the group A and group B.
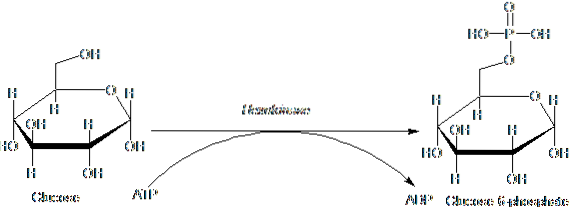
The enzymes belonging to the HK family are characterized by phosphorylating glucose at the expense of ATP as a phosphoryl group donor molecule, and its members differ among themselves mainly with respect to their molecular weight and substrate specificity..
The HK group includes the enzymes of eukaryotic organisms (ATP: D-hexose 6-phosphotransferases), while group A is represented by the enzymes of gram-negative bacteria, cyanobacteria, amitochondriate protists and trypanosomatids, and group B contains the enzymes gram-positive bacteria and crenoqueotic organisms.
The enzymes of groups A and B are also known as glucokinases (GlcKs), as they are capable of exclusively phosphorylating glucose, which is why these enzymes are called ATP: D-glucose 6-phosphotransferases.
As a glycolytic enzyme, hexokinase has great metabolic importance, since without it this important pathway would not be possible and cells highly dependent on carbohydrate consumption, such as, for example, the brain and muscle cells of many mammals, would have serious functional and physiological impediments in general.
Article index
As will be seen later, different types of hexokinase enzymes exist in mammals and other vertebrates (as well as in single-celled organisms such as yeast). In mammals, four have been described: isoforms I, II, III and IV.
The first three isozymes have 100 kDa molecular weight, but isozyme IV has 50 kDa. These isoenzymes (especially I-III) exhibit high sequence similarity between them with respect to their C- and N-termini, as well as with other members of the hexokinase family..
The N-terminal domain of these enzymes is considered the "regulatory" domain, while the catalytic activity is carried out by the C-terminal domain (mammalian HK II has active sites in both domains).
The N-terminal domain is linked to the C-terminal domain through an alpha helix, each is roughly 50 kDa in molecular weight and has a binding site for glucose.
The tertiary structure of these enzymes is essentially composed of β-folded sheets mixed with alpha helices, the proportion of which varies with respect to the enzyme and the species in question; the binding site for ATP, the other substrate for hexokinase, is usually composed of five β sheets and two alpha helices.
Hexokinase has a transcendental function in the carbohydrate metabolism of most living beings, since it catalyzes the first step of the glycolytic pathway, mediating the phosphorylation of glucose inside the cell..
This first step of glycolysis, which consists of the transfer of a phosphoryl group from ATP (donor) to glucose, yielding glucose 6-phosphate and ADP, is the first of the two energy investment steps in the form of ATP.
Furthermore, the reaction catalyzed by hexokinase is the step of "activation" of glucose for its subsequent processing and represents a step of "commitment", since the glucose thus phosphorylated cannot leave the cell by means of its conventional transporters in the membrane. plasmatic.
The product of the reaction catalyzed by hexokinase, that is, glucose 6-phosphate, is a branch point, since it is the first substrate used in the pentose phosphate pathway and in the synthesis of glycogen in many animals (and the starch in plants).
The function of hexokinase in plants is not very different from that of animals or microorganisms, however, in higher plants this enzyme also functions as a "sensor" of the concentration of sugars..
The importance of this function in these organisms has to do with the participation of sugars as regulatory factors in the expression of genes that are involved in different metabolic processes such as:
- Photosynthesis
- The glyoxylate cycle
- The breathing
- The degradation or synthesis of starch and sucrose
- Nitrogen metabolism
- Defense against pathogens
- Regulation of the cell cycle
- The healing response
- Pigmentation
- Senescence, among others.
This function of hexokinase as a "sensor" of the amount of intracellular glucose has also been described for yeast and mammals..
In nature there are different forms of hexokinases and this depends fundamentally on the species that is considered.
In man and other vertebrate animals, for example, the existence of 4 different isoforms of the hexokinase enzyme in the cytosolic compartment has been shown, which have been denoted with the Roman numerals I, II, III and IV.
Isozymes I, II and III have a molecular weight of 100 kDa, are inhibited by their reaction product (glucose 6-phosphate) and are very glucose-related, that is, they have a very low Km constant. However, these enzymes have poor substrate specificity, being able to phosphorylate other hexoses such as fructose and mannose..
Isoenzyme IV, also known as glucokinase (GlcK), has only 50 kDa in molecular weight and, despite being poorly related (high Km values), it has a high specificity for glucose as a substrate and is not subjected to them. regulatory mechanisms than the other three isoenzymes.
Glucokinase (isoenzyme IV of the hexokinase of many mammals) is mainly found in the liver and helps this organ in the "adjustment" of its rate of glucose consumption in response to variations of this substrate in the circulating blood..
The three genes that code for hexokinases I, II and III in animals appear to have the same 50 kDa ancestor that was duplicated and fused in the genome, which seems evident when it is observed that the catalytic activity of forms I and III resides only at the C-terminal end.



Yet No Comments