The histones are basic proteins that interact with DNA for the formation of nucleosomes, which make up the chromatin strands that make up chromosomes in eukaryotic organisms.
Nucleosomes, complexes made up of DNA and proteins, were discovered in 1974 and it is histones that assemble this basal level of chromatin organization. However, the existence of histone proteins has been known since before the 1960s..
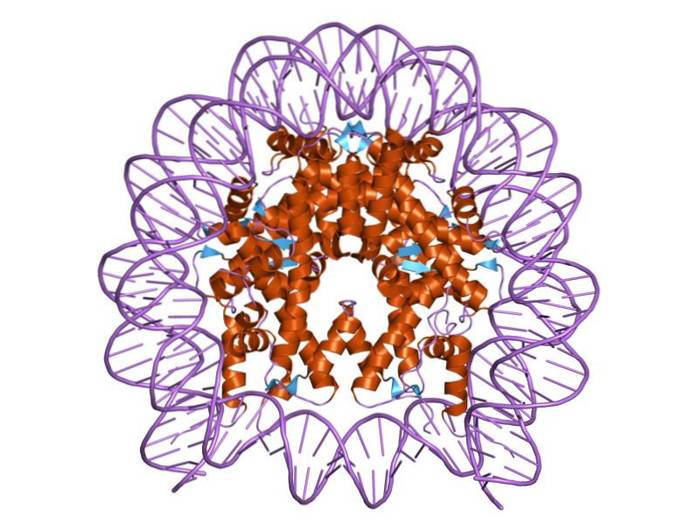
Histones are organized in such a way that the double-band DNA wraps around a protein center made up of these proteins that interact closely with each other. The histone center is disk-shaped and DNA goes around 1.7 times.
Multiple hydrogen bonds allow DNA to bind to the protein center formed by the histones in each nucleosome. These bonds are formed, for the most part, between the amino acid backbones of histones and the sugar-phosphate backbone of DNA. Some hydrophobic interactions and ionic bonds also participate.
Proteins known as "chromatin remodeling complexes" are responsible for the breakdown and formation of the binding bonds between DNA and histones, allowing the transcriptional machinery to enter the DNA contained in nucleosomes.
Despite the proximity of nucleic acids to the protein center formed by histones, these are arranged in such a way that, if necessary, they allow the entry of transcription factors and other proteins related to gene expression or gene silencing..
Histones can undergo various modifications that generate multiple variants, making possible the existence of many different forms of chromatin that have the property of modulating genetic expression in different ways..
Article index
They are one of the most conserved eukaryotic proteins in nature. For example, pea histone H4 has been shown to differ in only two of the 102 amino acid positions of the cow H4 protein..
Histones are relatively small proteins, with no more than 140 amino acids. They are rich in basic amino acid residues, so they have a net positive charge, which contributes to their interaction with the negatively charged nucleic acid to form nucleosomes..
Nucleosomal and bridging histones are known. The nucleosomal histones are H3, H4, H2A, and H2B, while the binding histones belong to the H1 family of histones..
During nucleosome assembly, specific dimers H3-H4 and H2A-H2B are initially formed. Two H3-H4 dimers then join together to form tetramers that later combine with the H2A-H2B dimers, forming the octameric center.
All histones are synthesized mainly during the S phase of the cell cycle, and nucleosomes are assembled in the nascent DNA helices, just after the replication fork..
The general structure of histones includes a basic amino acid region and a globular carboxyl region highly conserved among eukaryotic organisms..
A structural motif known as "histone fold", composed of three alpha helices connected by two hairpins and forming a small hydrophobic center, is responsible for the protein-protein interactions between the histones that make up the nucleosome..
It is this fold of histones that makes up the globular carboxyl domain of these nucleosomal proteins in all eukaryotes..
Histones also have small "tails" or amino-terminal and other carboxyl-terminal regions (accessible to proteases), no more than 40 amino acids long. Both regions are rich in basic amino acids that can undergo multiple post-translational covalent modifications.
In eukaryotes there are two families of binding histones, differentiated from each other by their structure. Some have a tripartite structure, with the globular domain described above flanked by "unstructured" N- and C-terminal domains; while others only have a C-terminal domain.
Although most histones are conserved, specific variants may arise during embryogenesis or maturation of specialized cells in some organisms. Some structural variations have to do with post-translational modifications such as the following:
-Phosphorylation: it is thought to be related to the modification of the degree of condensation of chromatin and it occurs commonly in serine residues.
-Acetylation: associated with chromosomal regions that are transcriptionally active. It normally occurs on the side chains of lysine residues. When occurring on these residues, their positive charge decreases, thereby reducing the affinity of proteins for DNA..
-Methylation: can occur as mono-, di- or trimethylation of lysine residues that protrude from the protein core.
Specific enzymes are responsible for these covalent modifications in histones. These enzymes include histone-acetyl transferases (HATs), histone-deacetylases complexes (HDACs), and histone-methyltransferases and demethylases..
The characterization of the histones has been carried out by various biochemical techniques, among which the chromatographies that are based on weak cation exchange resins stand out..
Certain authors establish a form of classification in which 5 main types of histones are distinguished in eukaryotes: FI, with 21 kDa proteins; F2A1 or FIV, of plus or minus 11.3 kDa; F2A2 or FIIbI, 14.5 kDa; F2B or FIIb2, with a molecular weight of 13.7 kDa, and F3 or FIII, of 15.3 kDa.
All these types of histones, with the exception of the IF group, are found in equimolar amounts in cells..
Another classification, with the same validity and perhaps the most widely used today, proposes the existence of two different types of histones, namely: those that are part of the octamer of the nucleosome and the binding or bridging histones, which join the nucleosomes between Yes.
Some variants can also occur between species and unlike core histones, variants are synthesized during interphase and are inserted into preformed chromatin through a process dependent on the energy released from ATP hydrolysis..
The center of a nucleosome consists of a pair of each of the four constituent histones: H2a, H2b, H3, and H4; on which DNA segments of about 145 base pairs are wound.
Histones H4 and H2B are, in principle, invariant. Some variations are evident, however, in histones H3 and H2A, whose biophysical and biochemical properties alter the normal nature of the nucleosome..
A variant of histone H2A in humans, the H2A.Z protein has a large acidic region and may promote nucleosome stability depending on the histone H3 variants with which it is associated..
These histones show some variability between species, a special case being that of histone H2B, for which the first third of the molecule is highly variable..
The binding or bridging histones are class H1 histones. These are responsible for the union between nucleosomes and the protection of the DNA that protrudes at the beginning and end of each particle..
Unlike nucleosomal histones, not all H1-type histones possess the globular region of the histone "fold". These proteins bind to DNA between nucleosomes, facilitating a shift in chromatin balance towards a more condensed and less active state, transcriptionally speaking..
Studies have linked these histones to aging, DNA repair, and apoptotic processes, which is why they are thought to play a crucial role in maintaining genomic integrity..
All the amino acid residues of histones participate, in one way or another, in their interaction with DNA, which explains the fact that they are so conserved among the kingdoms of eukaryotic organisms.
The participation of histones in the packaging of DNA in the form of chromatin is of great relevance for complex multicellular organisms in which the different cell lines can specialize only by changing the accessibility of their genes to the transcriptional machinery..
The transcriptionally active genomic regions are dense in nucleosomes, suggesting that the association of DNA with histone proteins is crucial for the negative or positive regulation of their transcription..
Likewise, throughout the life of a cell, the response to a large number of stimuli, both internal and external, depends on small changes in chromatin, which normally have to do with remodeling and post-translational modification of the histones found in close relationship with DNA.
Multiple histone variables serve different functions in eukaryotes. One of them has to do with the participation of a variant of histone H3 in the formation of the centromeric structures responsible for the segregation of chromosomes during mitosis..
The counterpart of this protein in other eukaryotes has been shown to be essential for the assembly of the protein kinetochore to which spindle microtubules bind during mitosis and meiosis..



Yet No Comments