The nucleosome It is the basic unit of DNA packaging in eukaryotic organisms. It is therefore the smallest chromatin compression element.
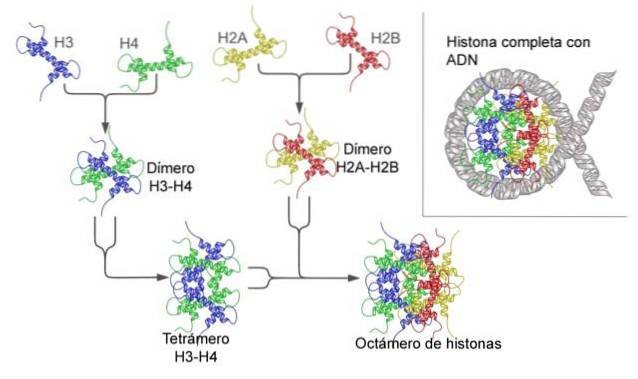
The nucleosome is built as an octamer of proteins called histones, or a drum-shaped structure on which about 140 nt of DNA is wound, making almost two complete turns..
Additionally, an additional 40-80 nt of DNA is considered to be part of the nucleosome, and it is the DNA fraction that allows physical continuity between one nucleosome and another in more complex chromatin structures (such as the 30 nm chromatin fiber)..
The histone code was one of the first molecularly best understood epigenetic control elements.
Article index
Nucleosomes allow:
In its most basic sense, nucleosomes are made up of DNA and proteins. DNA can be virtually any double-band DNA present in the nucleus of the eukaryotic cell, while nucleosomal proteins all belong to the set of proteins called histones..
Histones are small proteins with a high load of basic amino acid residues; This makes it possible to counteract the high negative charge of DNA and establish an efficient physical interaction between the two molecules without reaching the rigidity of the covalent chemical bond..
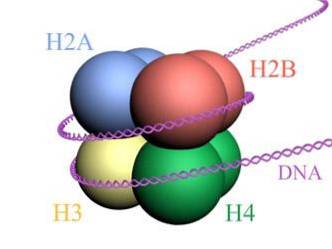
Histones form a drum-like octamer with two copies or monomers of each of histones H2A, H2B, H3, and H4. The DNA makes almost two complete turns on the sides of the octamer and then continues with a fraction of linker DNA that associates with histone H1, to return to give two complete turns in another histone octamer.
The octamer set, associated DNA, and its corresponding linker DNA, is a nucleosome.
Genomic DNA is made up of extremely long molecules (more than a meter in the case of humans, considering all their chromosomes), which must be compacted and organized within an extremely small nucleus.
The first step in this compaction is carried out through the formation of nucleosomes. With this step alone, DNA is compacted about 75 times.
This gives rise to a linear fiber from which subsequent levels of chromatin compaction are built: the 30 nm fiber, the loops, and the loops of loops..
When a cell divides, either by mitosis or meiosis, the ultimate degree of compaction is the mitotic or meiotic chromosome itself, respectively..
The fact that histone octamers and DNA interact electrostatically explains in part their effective association, without losing the fluidity required to make nucleosomes dynamic elements for compacting and decompacting chromatin..
But there is an even more surprising interaction element: the N-terminal ends of the histones are exposed outside the interior of the octamer, which is more compact and inert..
These ends not only physically interact with the DNA, but also undergo a series of covalent modifications on which the degree of compaction of the chromatin and the expression of the associated DNA will depend..
The set of covalent modifications, in terms of type and number, among other things, is collectively known as the histone code. These modifications include phosphorylation, methylation, acetylation, ubiquitination, and sumoylation of arginine and lysine residues at the N-termini of histones..
Each change, together with others within the same molecule or in residues of other histones, particularly histones H3, will determine the expression or not of the associated DNA, as well as the degree of compaction of the chromatin..
As a general rule, it has been seen, for example, that hypermethylated and hypoacetylated histones determine that the associated DNA is not expressed and that chromatin is present in a more compact state (heterochromatic, and therefore inactive)..
In contrast, euchromatic DNA (less compact, and genetically active) is associated with a chromatin whose histones are hyperacetylated and hypomethylated..
We have already seen that the covalent modification status of histones can determine the degree of expression and local chromatin compaction. At global levels, chromatin compaction is also being regulated by covalent modifications of histones in nucleosomes.
For example, it has been shown that constitutive heterochromatin (which is never expressed, and is densely packed) tends to be attached to the nuclear lamina, leaving the nuclear pores free..
For its part, constitutive euchromatin (which is always expressed, such as that which includes cell maintenance genes, and is located in regions of lax chromatin), does so in large loops that expose the DNA to be transcribed to the transcription machinery.
Other regions of genomic DNA oscillate between these two states depending on the development time of the organism, growth conditions, cell identity, etc..
In order to fulfill their plan for cell development, expression and maintenance, the genomes of eukaryotic organisms must finely regulate when and how their genetic potentialities must manifest..
Starting then from the information stored in their genes, these are located in the nucleus in particular regions that determine their transcriptional state..
We can say, therefore, that another of the fundamental roles of nucleosomes, through the changes in chromatin that it helps to define, is the organization or architecture of the nucleus that houses them..
This architecture is inherited and is phylogenetically conserved thanks to the existence of these modular elements of informational packaging.



Yet No Comments