The parthenogenesis is the production of an embryo from a female gamete without genetic contribution from a male gamete, with or without eventual development into an adult. In most cases there is no participation of the male gamete in the reproductive process.
However, there is a particular case of parthenogenesis called gymnogenesis, in which their participation is necessary. In this case, the sperm enters the egg and activates it to start the development of a new organism..

Parthenogenesis is a very common phenomenon among plants and animals. There are estimates that it can occur in up to 1% of the total number of known species.
It is a mode of reproduction that can occur in practically all large groups of animals and plants. Perhaps the exception is in the more evolved taxa, such as gymnosperms and mammals, in which there are no reliable records of its occurrence..
Article index
The simplest concept of parthenogenesis indicates that it is the development of the egg cell in a new individual without fertilization occurring. However, in many animals the embryos produced without fertilization suffer a high mortality.
In some other cases, the concurrence of a male gamete is necessary just to activate development. For this reason it can be said that parthenogenesis consists of "the production of an embryo from a female gamete without any genetic contribution from a male gamete with or without eventual development in an adult".
Depending on the cytological mechanisms involved, parthenogenesis can be of several types, including:
Also called facultative parthenogenesis, it is called in this way when the development of unfertilized eggs occurs occasionally and spontaneously. This type of parthenogenesis is very common in animals.
According to some authors, it is the only real type of parthenogenesis. Gamete formation in aticoparthenogenesis may or may not involve meiotic divisions. Depending on the presence or absence of meiosis, this parthenogenesis can in turn be divided into:
Also called ameiotic or diploid. In this there is suppression of meiosis. The offspring develop from unfertilized eggs, through mitotic division.
It gives rise to organisms that are genetically identical to the mother. This type of parthenogenesis usually occurs in rotifers and most groups of arthropods..
Also called meiotic or haploid. In this process, meiosis is maintained. The restoration of the diploid condition occurs by the duplication or fusion of gametes produced by the mother. This type of parthenogenesis occurs very frequently in insects.
Gynogenesis is a special type of sexual reproduction. In this, it is necessary for the sperm to penetrate the ovum to activate embryonic development..
But, contrary to normal fertilization, fusion of the male and female nuclei does not occur. After the fusion of gametes, the sperm chromosomes degenerate within the cytoplasm of the egg or can be expelled from the zygote.
The gynogenetic embryo will develop only at the expense of the nucleus of the ovum. Because of this, the gynogenetic pups are all identical females to the mother..
This type of reproduction can occur when gynogenetic females mate with bisexual males of the same or related species. Some authors do not consider it a proper parthenogenesis.
It is a "hemiclonal" mode of reproduction. In this, parents of different species mate and produce hybrids. Half of the genome is transmitted sexually, while the other half is "clonally".
Sperm fuse in the nucleus of the ovum, and paternal genes are expressed in somatic tissues, but are systematically excluded from the germ line. Only the mother will pass on the genome to the next generation.
This type of parthenogenesis usually occurs in fish species of the genus Poeciliopsis, and has also been observed in the desert ant Cataglyphis hispanica.
Some authors prefer a more utilitarian classification of this type of reproduction, differentiating parthenogenesis into two other types:
It is characterized by the coexistence of a bisexual and a parthenogenetic form, in a single species or in species that are phylogenetically close, but have different geographical distribution..
Parthenogenetic organisms tend to occupy different ranges from their close relatives who reproduce sexually. Asexual organisms tend to have higher distributions latitudinally or altitudinally, on islands, in xerophilic environments, or in disturbed habitats..
This type of parthenogenesis has been observed in some species of plants, worms, crustaceans, insects and lizards..
Organisms can reproduce both sexually and parthenogenetically. During some periods of the year only females are produced by parthenogenesis.
However, in other periods the females will produce both females and males who will reproduce sexually..
In bisexual species where offspring are produced by parthenogenesis, they will generally produce parthenogenetic females. These emerging unisexual lineages can differ greatly phenotypically and genotypically from their bisexual congeners. There are several mechanisms that can give rise to these parthenogenetic lineages.
Loss of sexual interaction occurs through mutations in genes that suppress meiosis, modify the induction of sex by environmental conditions, and regulate hormonal expression.
In extreme cases, the mutation could act by "repairing" the genotype of a strictly parthenogenetic lineage, which could produce parthenogenetic males and females..
Hybridization is the most common way to produce parthenogenetic lineages in animals, and it can be observed in snails, insects, crustaceans and most unisexual vertebrates..
They originate from the crossing of two bisexual species that have high heterozygosity and typical alleles of parental species. In these, meiosis can be an impediment, causing the loss of sexuality.
It occurs by hybridization between parthenogenetic females and males of the same or closely related species. It is believed to be the main cause of polyploidy in unisexual organisms..
The flow of genes between sexual and parthenogenetic lineages allows the spread of genes in a contagious way. Due to this, sexual organisms can originate in turn, or create a new parthenogenetic line.
Wolbachia pipientis is a species of bacteria of the phylum Proteobacteria that is home to about 20% of all insect species.
It is responsible for reproductive manipulations in its hosts, such as cytoplasmic incompatibility, feminization of genetic males, male death, and parthenogenesis. Infects arthropods and nematodes.
It is transmitted parentally. This bacterium is capable of inducing parthenogenesis in parasitoid wasps of the genus Trichogramma as well as mites and other arthropods.
On the other hand, Xiphinematobacter, other bacteria, affects Dorylaimida nematodes, also causing parthenogenesis.
In many species, parthenogenetic lineages are generated by a single mechanism. However, in other species they can arise through various mechanisms. For example, the parthenogenetic lineages of ostracods often have a dual origin.
Diploid clones originate from spontaneous loss of sexuality, while polyploid clones arise from hybridization between parthenogenetic males and females of the same or related species.
Another example is the case of the aphid Rhopalosiphum padi. In this species, parthenogenetic lineages can arise from three different origins: spontaneous, hybrid or contagious..
Among the Rotifera there are species that reproduce only by female apomictic parthenogenesis and species that alternate this parthenogenesis with ordinary sexual reproduction.
The transition between asexual and sexual reproduction is controlled by the environment. The success of rotifer species that have completely lost sexual reproduction is due, according to some authors, to the accumulation of mutations during periods of exponential apomictic parthenogenetic reproduction.
This, together with the "mitotic" crossing, would allow sufficient genotypic diversity to be produced to adapt to different environmental conditions. In this way, a great advantage of sexual reproduction would be eliminated..
Parthenogenesis has been reported for some species of gastropod mollusks. Among these species are Potamopyrgus antipodarum, Tarebia granifera, and all species of the genus Melanoids.
All representatives of the latter genus, except the diploid race of M. tuberculata, they are polyploid.
This type of reproduction has been documented for numerous groups of crustaceans, including notostracos, conchostracos, anostracos, cladocerans, decapods and ostracods..
In Cladocera the typical form of reproduction is by cyclic parthenogenesis. Females reproduce parthenogenetically from spring to summer.
When environmental conditions are adverse, organisms reproduce sexually, in order to form encyst eggs that can survive long periods in dormancy states..
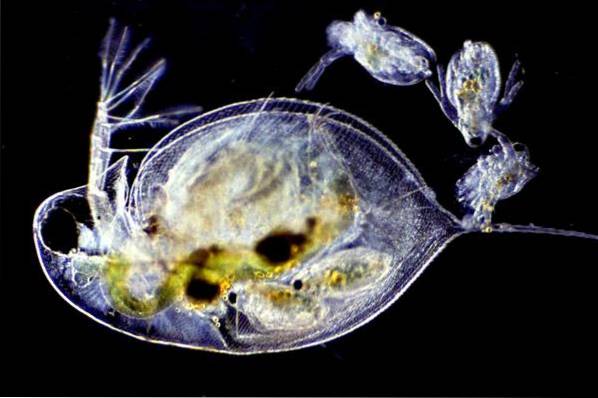
The marble crabs (Procambarus fallax shape virginalis) are the only known decapod crustaceans that reproduce only by parthenogenesis.
Among cartilaginous fish, parthenogenesis occurs at least in the eagle ray, zebra shark, and hammerhead shark. In bony fish, hybridgenesis has been reported for species of the genus Poecilliopsis.
Some other fish can alternate sexual and parthenogenetic reproduction. Many species of lizards reproduce by parthenogenesis. It is believed that hybridization is the main cause of this type of reproduction in the same.
Ticoparthenogenesis has also been reported in other groups of reptiles, mainly pythons and other snakes. In birds, spontaneous parthenogenesis has been observed in chickens, turkeys and some species of quail..
In mammals, maternal and paternal genomes are necessary for normal embryonic development. Because of this, parthenogenesis does not occur naturally in these organisms..
This has been achieved experimentally in the laboratory. However, induced parthenogenesis often results in abnormal development..
Many plant species present well-defined patterns of geographic parthenogenesis, where the parthenogenetic forms are located more towards the cold zones. The sexual forms, meanwhile, are more tropical than their asexual peers..

Yet No Comments