The uracil It is a pyrimidine-type nitrogenous base, found in ribonucleic acid (RNA). This is one of the characteristics that differentiate RNA from deoxyribonucleic acid (DNA), since the latter has thymine instead of uracil. Both substances, uracil and thymine, only differ in that the second has a methyl group.
From an evolutionary point of view, it has been proposed that RNA was the first molecule to store genetic information and function as a catalyst in cells, before DNA and enzymes. Because of this, uracil is thought to have played a key role in the evolution of life.
In living things, uracil is not found in a free form, but commonly forms nucleotides monophosphate (UMP), diphosphate (UDP), and triphosphate (UTP). These uracil nucleotides have different functions, such as RNA and glycogen biosynthesis, isomeric interconversion of sugars, and regulation of glutamine synthase..
Article index
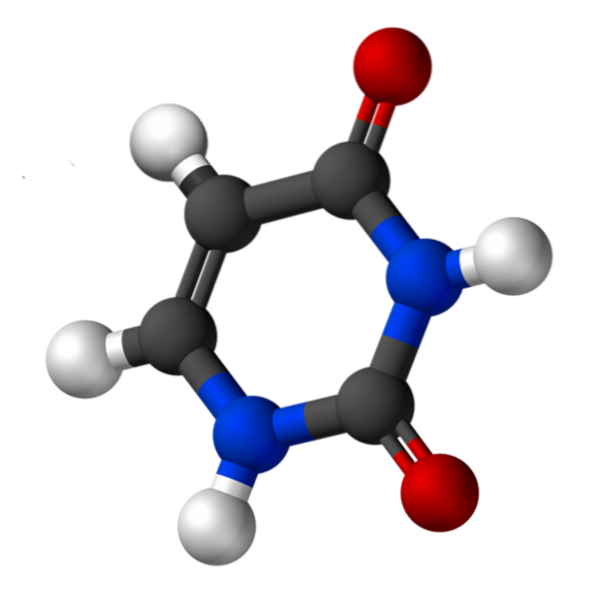
Uracil, called 2,4-dioxypyridine, has the empirical formula C4H4NtwoORtwo, whose molecular weight is 112.09 g / mol, and is purified as a white powder.
The structure of uridine is a heterocyclic ring with four carbon atoms and two nitrogen atoms, with alternating double bonds. Is planar.
It has a solubility of 50mg / ml, at 25ºC, in 1M sodium hydroxide, and a pKa between 7.9 and 8.2. The wavelength where its maximum absorbance occurs (ʎmax) is between 258 and 260 nm.
There is a common pathway for pyrimidine nucleotide biosynthesis (uracil and cytokine). The first step is the biosynthesis of carbamoyl phosphate from COtwo and NH4+, which is catalyzed by carbamoyl phosphate synthetase.
Pyrimidine is built from carboyl phosphate and aspartate. Both substances react and form N-carbamoylaspartate, a reaction catalyzed by aspartate transcabamoylase (ATCase). The closure of the pyrimidine ring is caused by dehydration catalyzed by dihydrootase, and produces L-dihydrorotate..
L-dihydrorotate is oxidized and converted to orotate; electron acceptor is NAD+. It is a reaction catalyzed by dihydroorotate dehydrogenase. The next step is the transfer of the phosphoribosyl group, from phosphoribosyl pyrophosphate (PRPP), to orotate. Forms orotidylate (OMP) and inorganic pyrophosphate (PPi), catalyzed by orotate phosphoribosyl transferase.
The last step consists of the decarboxylation of the pyrimidine ring of the orotidylate (OMP). Forms uridylate (uridine-5'-monophosphate, UMP), which is catalyzed by a decarboxylase.
Then, through the participation of a kinase, a phosphate group is transferred from ATP to UMP, forming UDP (uridine-5'-diphosphate). The latter is repeated, forming UTP (uridin-5'-triphosphate).
In bacteria, regulation of pyrimidine biosynthesis occurs through negative feedback, at the level of aspartate transcabamoylase (ATCase).
This enzyme is inhibited by CTP (cytidine-5'-triphosphate), which is the end product of the pyrimidine biosynthetic pathway. ATCase has regulatory subunits that bind to the allosteric regulator CTP.
In animals, the regulation of pyrimidine biosynthesis occurs through negative feedback, at the level of two enzymes: 1) carbamoyl phosphate synthase II, which is inhibited by UTP and activated by ATP and PRPP; and 2) OMP decarboxylase, which is inhibited by the product of the reaction that it catalyzes, UMP. The rate of biosynthesis of OMP varies with the availability of PRPP.
Uracil is present in all types of RNA, such as messenger RNA (mRNA), transfer RNA (tRNA), and ribosomal RNA (rRNA). The biosynthesis of these molecules occurs through a process called transcription..
During transcription, the information contained in DNA is copied into RNA by an RNA polymerase. The reverse process, in which the information contained in RNA is copied into DNA, occurs in some viruses and plants through reverse transcriptase..
RNA biosynthesis requires nucleoside triphosphate (NTP), namely: uridine triphosphate (UTP), cytidine triphosphate (CTP), adenine triphosphate (ATP) and guanine triphosphate (GTP). The reaction is:
(RNA)n residues + NTP -> (RNA)n + 1 residue + PPi
Hydrolysis of inorganic pyrophosphate (PPi) provides the energy for RNA biosynthesis.
Sugar esters are very common in living organisms. Some of these esters are the nucleoside ester diphosphates, such as UDP-sugars, which are very abundant in cells. UDP-sugars participate in the biosynthesis of disaccharides, oligosaccharides and polysaccharides.
In plants, sucrose biosynthesis occurs through two pathways: a primary and a secondary.
The main pathway is the transfer of D-glucose from UDP-D-glucose to D-fructose to form sucrose and UDP. The secondary pathway includes two steps: it begins with UDP-D-glucose and fructose-6-phosphate and ends with the formation of sucrose and phosphate..
In the mammary glands, lactose biosynthesis occurs from UDP-D-galactose and glucose.
In plants, cellulose biosynthesis is carried out by the continuous condensation of beta-D-glucosyl residues, from UDP-glucose to the non-reducing end of the growing polyglucose chain. Similarly, amylose and amylopectin biosynthesis requires UDP-glucose as a glucose donor substrate to the growing chain..
In animals, both UDP-glucose and ADP-glucose are used for glycogen biosynthesis. Similarly, chondroitin sulfate biosynthesis requires UDP-xylose, UDP-galactose, and UDP-glucuronate..
The conversion of galactose to a glycolysis intermediate occurs through the Leloir pathway. One of the steps in this pathway is catalyzed by the enzyme UDP-galactose-4-epimerase, which facilitates the interconversion of UDP-galactose to UDP-glucose..
During glycoprotein biosynthesis, proteins cross the cis, middle, and trans sacs of the Golgi apparatus.
Each of these sacs has a set of enzymes that process glycoproteins. Sugar monomers, such as glucose and galactose, are added to the protein oligosaccharide from UDP-hexose and other nucleotides-hexose.
The hexose nucleotides are transported to the Golgi cisterns by antiport. UDP-galactose (UDP-Gal) and UDP-N-acetylgalactosamine (UDP-GalNAc) enter the cisterns from the cytosol by exchange for UMP.
In the Golgi cistern, a phosphatase hydrolyzes a phosphate group on UDP and forms UMP and Pi. UDP comes from the reactions catalyzed by galactosyltransferase and N-acetylgalactosamyltransferase. UMP formed by phosphatase serves for nucleotide-hexose exchange.
A regulatory mechanism of glutamine synthase is covalent modification, which consists of adenylation, which inactivates it, and dedenylation, which activates it. This covalent modification is reversible and catalyzed by adenyltransferase..
Adenyltransferase activity is modulated by the binding of the PII protein, which is regulated by a covalent modification, uridinylation.
Both uridylation and deuridylation are carried out by uridylyltransferase. In this enzyme, uridylation activity is due to glutamine and phosphate, and is activated by binding of alpha-ketoglutarate and ATP to PII.
Some mRNAs are edited before translation. In some eukaryotic organisms, such as Trypanosoma brucei, there is RNA editing of the cytochrome oxidase subunit II gene transcript. This happens through the insertion of uracil residues, a reaction catalyzed by the terminal uridyltransferase..
A guide RNA, complementary to the edited product, acts as a template for the editing process. The base pairs formed between the initial transcript and the guide RNA implies G = U base pairs that are not Watson-Crick and are common in RNA..
Under physiological conditions, the biosynthesis of glycogen from glucose-1-phosphate is thermodynamically impossible (ΔG positive). Due to this, prior to biosynthesis, the activation of glucose-1-phosphate (G1P) occurs. This reaction combines G1P and UTP to form uridine diphosphate glucose (UDP-glucose or UDPG)..
The reaction is catalyzed by UDP-glucose pyrophosphorylase, and is as follows:
G1P + UTP -> UDP-glucose + 2Pi.
The Gibbs free energy variation in this step is large and negative (-33.5 KJ / mol). During the reaction to oxygen, G1P attacks the alpha phosphorus atom of UTP and forms UDP-glucose and inorganic pyrophosphate (PPi). Next, the PPi is hydrolyzed by an inorganic pyrophosphatase, whose hydrolysis energy is what drives the general reaction.
UDP-glucose is a "high energy" substance. It allows to form the glycosidic bonds between the glucose residue and the growing polysaccharide chain. This same energetic principle is applicable to reactions where UDP-sugars participate, such as the biosynthesis of disaccharides, oligosaccharides and glycoproteins..
There are DNA lesions that occur spontaneously. One of these lesions is the spontaneous deamination of cytokine, and its consequent conversion to uracil. In this case, repair takes place by removing the modified DNA base by an enzyme called uracil DNA glycosylase..
The enzyme uracil DNA glycosylase removes the damaged cytokine (uracil), producing a deoxyribose residue that lacks the nitrogen base, called the AP site (apurinic-apyrimidinic site).
The enzyme AP endonuclease then cuts the phosphodiester backbone of the AP site, removing the sugar-phosphate residue. DNA polymerase I restores the damaged strand.


Yet No Comments