The zinc fingers (ZF) are structural motifs present in a large number of eukaryotic proteins. They belong to the group of metalloproteins, as they are capable of binding the zinc metal ion, which they require for their operation. More than 1,500 ZF domains are predicted to exist in around 1,000 different proteins in humans.
The term zinc finger or "zinc finger" was first coined in 1985 by Miller, McLachlan and Klug, while studying in detail the small DNA-binding domains of the transcriptional factor TFIIIA of Xenopus laevis, described by other authors some years before.
Proteins with ZF motifs are among the most abundant in the genome of eukaryotic organisms and participate in a variety of essential cellular processes, including gene transcription, protein translation, metabolism, the folding and assembly of other proteins and lipids. , programmed cell death, among others.
Article index
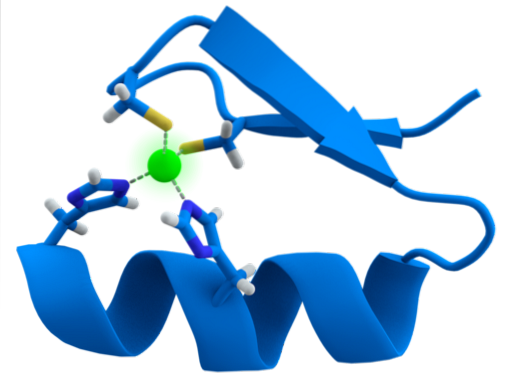
The structure of the ZF motifs is extremely conserved. Usually these repeated regions have 30 to 60 amino acids, whose secondary structure is found as two antiparallel beta sheets that form a hairpin and an alpha helix, which is denoted as ββα.
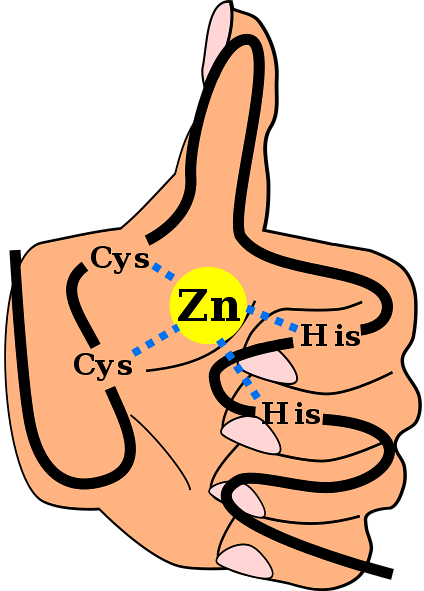
Said secondary structure is stabilized by hydrophobic interactions and by the coordination of a zinc atom given by two cysteine and two histidine residues (CystwoHistwo). However, there are ZFs that can coordinate more than one zinc atom and others where the order of the Cys and His residues varies..
The ZFs can be found repeated in batches, linearly configured in the same protein. They all have similar structures, but they can be chemically differentiated from each other by variations of key amino acid residues for the fulfillment of their functions..
A common feature among ZFs is their ability to recognize DNA or RNA molecules of different lengths, which is why they were initially considered only as transcriptional factors.
In general, recognition is of 3bp regions in DNA and is achieved when the protein with the ZF domain presents the alpha helix to the major groove of the DNA molecule.
There are different ZF motifs that differ from each other by their nature and the different spatial configurations achieved by the coordination bonds with the zinc atom. One of the classifications is as follows:
This is a commonly found motif in ZFs. Most C motifstwoHtwo they are specific for interaction with DNA and RNA, however, they have been observed to participate in protein-protein interactions. They have between 25 and 30 amino acid residues and are found within the largest family of regulatory proteins in mammalian cells.
They interact with RNA and some other proteins. They are mainly observed as part of some retrovirus capsid proteins, assisting in the packaging of viral RNA just after replication..
Proteins with this motif are enzymes responsible for DNA replication and transcription. A good example of these can be the coarse enzymes of phages T4 and T7.
This ZF family comprises transcription factors that regulate the expression of important genes in numerous tissues during cell development. Factors GATA-2 and 3, for example, are involved in hematopoiesis.
These domains are unique to yeast, specifically the GAL4 protein, which activates the transcription of genes involved in the use of galactose and melibiose..
These particular structures possess 2 subtypes of ZF domains (C3HC4 and C3HtwoC3) and are present in numerous animal and plant proteins.
They are found in proteins like RAD5, involved in DNA repair in eukaryotic organisms. They are also found in RAG1, essential for the reconfiguration of immunoglobulins.
This domain of ZF is highly conserved in the integrases of retroviruses and retrotransposons; binding to the target protein causes a conformational change in it.
Proteins with ZF domains serve a variety of purposes: they can be found on ribosomal proteins or on transcriptional adapters. They have also been detected as an integral part of the structure of yeast RNA polymerase II..
They appear to be involved in intracellular zinc homeostasis and in the regulation of apoptosis or programmed cell death. In addition, there are some proteins with ZF that function as chaperones for the folding or transport of other proteins..
Binding to lipids and a fundamental role in protein-protein interactions are also prominent functions of the ZF domains in some proteins..
Over the years, the structural and functional understanding of the ZF domains has allowed great scientific advances that involve the use of their characteristics for biotechnological purposes..
Since some proteins with ZF have a high specificity for certain DNA domains, much effort is currently invested in the design of specific ZFs, which can provide valuable advances in gene therapy in humans.
Interesting biotechnological applications also arise from the design of proteins with genetically engineered ZFs. Depending on the desired purpose, some of these can be modified by the addition of "poly zinc" finger peptides, which are capable of recognizing virtually any DNA sequence with high affinity and specificity..
Nuclease-modified genomic editing is one of the most promising applications today. This type of editing offers the possibility to carry out studies on genetic function directly in the model system of interest..
Genetic engineering using modified ZF nucleases has captured the attention of scientists in the field of genetic improvement of cultivars of plants of agronomic importance. These nucleases have been used to correct an endogenous gene that produces herbicide resistant forms in tobacco plants..
Nucleases with ZF have also been used for the addition of genes in mammalian cells. The proteins in question were used to generate a set of isogenic mouse cells with a series of alleles defined for an endogenous gene..
Such a process has a direct application in the labeling and creation of new allelic forms to study the relationships of structure and function in native conditions of expression and in isogenic environments..

Yet No Comments