
The innervation of the heart it is organized into sympathetic and parasympathetic innervation. Like any other organ, the heart receives an innervation whose fibers are classified as belonging to the autonomic nervous system (ANS), one of the two divisions of the peripheral nervous system and responsible for mediating sensitivity and controlling the organism's visceral activity.
Despite being a striated muscular organ, very similar to skeletal muscle, the heart does not receive innervation from the other division of the peripheral system that mediates somatic sensitivity and the activity of the muscles that causes joint displacements.
Any contractile process in skeletal muscle requires excitation induced by a somatic motor nerve fiber. The heart, for its part, does not need to be excited by anything external to itself, since it has the ability to spontaneously generate its own excitations.
Thus, one of the outstanding characteristics of cardiac autonomic innervation is represented by the fact that it is not a determinant of the contractile activity of the heart, which can continue after denervation, but rather exerts a modulating function of it..
Article index
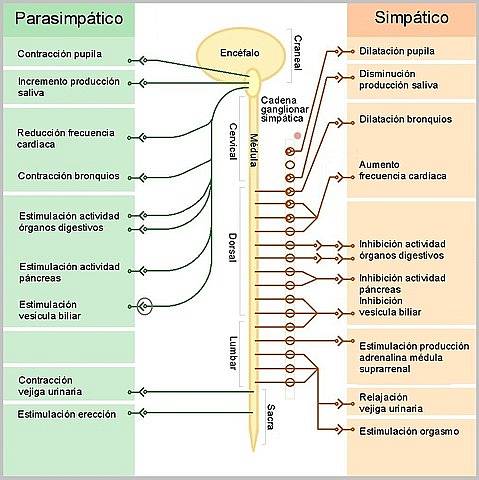
The efferent or motor portion of the autonomic nervous system is organized into two components: the sympathetic and the parasympathetic, systems made up of pathways that connect neurons in the central nervous system with visceral effector cells of the body on which they exert antagonistic effects..
Each of these pathways is a chain of two neurons:
The sympathetic preganglionic cells destined for the heart originate in cell conglomerates located in the lateral horns of the spinal cord, in the thoracic segments T1-T5. Cell conglomerates that together constitute a "spinal cardioaccelerator sympathetic center".
Its axons represent preganglionic fibers that are directed to the sympathetic ganglionic chain; especially to the superior, middle and inferior cervical ganglia, where they connect with the post ganglion neurons, whose axons are distributed with the superior, middle and inferior cardiac nerves.
Of these three nerves, the middle seems to be the one that exerts the greatest influence on cardiac functions, because the upper one is destined to the large arteries at the base of the heart and the lower one seems to conduct sensory or afferent information.
One more detail of the organization of cardiac sympathetic innervation is that the right sympathetic fibers seem to end mainly on the sinoatrial node, while the left ones influence the atrioventricular node, the conduction system and the contractile myocardium..
The sympathetic nervous system exerts a positive action on all cardiac functions, increasing the heart rate (chronotropism +), the force of contraction (inotropism +), the conduction of excitation (dromotropism +) and the relaxation rate (lusotropism +).
All these actions are exerted by the release of norepinephrine (NA) at the level of the sympathetic postganglionic endings on the cells of the cardiac nodes, the conduction system or on the atrial and ventricular contractile myocytes..
The actions of norepinephrine are triggered when this neurotransmitter binds to β1-type adrenergic receptors located on the membranes of cardiac cells and coupled to a Gs protein. This is a protein with three subunits (αsβγ) that when inactive has GDP bound to its αs subunit.
The norepinephrine-β1 receptor interaction causes the αs subunit to release its GDP and exchange it for GTP; In doing so, it separates from the βγ component and activates the membrane enzyme adenyl cyclase, which produces cyclic adenosine monophosphate (cAMP) as a second messenger that activates protein kinase A (PKA).
The phosphorylating activity of PKA is ultimately responsible for all the stimulatory actions that sympathetic fibers exert on the heart, and includes the phosphorylation of Ca ++ channels, troponin I and phospholamban.
The action on Ca ++ channels favors increases in heart rate, contractile force and conduction speed. The effects on troponin I and on phospholamban accelerate the relaxation process of the heart muscle.
The phosphorylation of troponin I causes this protein to accelerate the release process of Ca ++ from troponin C so that relaxation occurs faster. Phospholamban naturally inhibits the pump that reintroduces Ca ++ to the sarcoplasmic reticulum to terminate the contraction, an inhibition that is reduced when it is phosphorylated.
The parasympathetic innervation of the heart runs through the vagus nerve and its components have an organization of bineuronal chains similar to those of the sympathetic, with preganglionic neurons whose bodies are located in the dorsal motor nucleus of the vagus in the bulb, on the floor of the fourth ventricle..
Due to the reducing effects of cardiac activity that these neurons exert on the heart, they have been collectively called the "bulbar cardioinhibitor center". Its fibers separate from the vagal trunk in the neck and then intermingle with the cardiac sympathetic fibers to form a plexus..
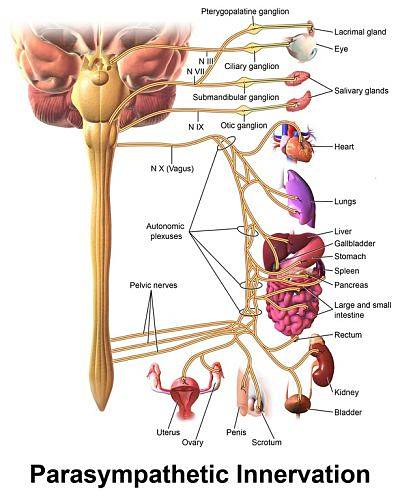
The parasympathetic ganglia are found in the vicinity of the heart and the postganglionic fibers on the right side mainly terminate in the sinoatrial node, the natural pacemaker of the heart, and the left in the atrioventricular node and in the atrial contractile myocytes.
Parasympathetic activity directed at the heart results in a negative effect on some cardiac functions such as a decrease in frequency (inotropism -), in the conduction speed in the AV node (dromotropism -) and a reduction in the contractile force of the atria (inotropism earphone -).
The scant or even non-existent innervation of the parasympathetic on the ventricular myocardium rules out a negative inotropic effect of this autonomic division on the contractile force of this muscle.
The aforementioned vagal actions on the heart are exerted by the release of acetylcholine (ACh) at the level of the parasympathetic postganglionic endings on the cells of the cardiac nodes and the atrial contractile myocytes..
The actions of acetylcholine are triggered when it binds to muscarinic cholinergic receptors type M2 located on the membranes of the mentioned cells and coupled to a Gi protein. It has three subunits (αiβγ) and when it is inactive it has GDP attached to its αi subunit..
The acetylcholine-M2 receptor interaction releases the αi subunit. This inhibits adenyl cyclase, less cAMP is produced and the activity of PKA and phosphorylation of Ca ++ channels are reduced, effects contrary to those of NA released by the sympathetic. The βγ component activates a K + current (IKACh).
The reduction of the phosphorylation of Ca ++ channels reduces the depolarizing current of this ion, while the appearance of the IKACh current introduces a hyperpolarizing current that opposes the spontaneous depolarization that produces the action potentials (AP) in nodular cells..
The reduction in the depolarizing Ca ++ current combined with the increase in the hyperpolarizing K + current slows the spontaneous depolarization process that automatically brings the membrane potential to the threshold level at which the action potential is triggered..
This effect can be of such magnitude that an intense stimulation of the vagus nerve can stop the heart, due to the disappearance of the action potentials of the pacemaker cells or due to a total blockage of the atrioventricular node that does not allow the potentials to pass. action from the right atrium to the ventricles.
Both the sympathetic and the parasympathetic are always active, exerting a permanent tonic action on the heart, so that cardiac functions at rest are the result of spontaneous cardiac activity tonically modulated by these two antagonistic influences..
The parasympathetic tone is greater than the sympathetic tone, which is deduced from the fact that when the heart is surgically or pharmacologically “denervated”, it accelerates, increasing the heart rate.
The increased metabolic demands of the body require an increase in cardiac activity that is achieved automatically by increasing the action that the sympathetic exerts on the heart and reducing the parasympathetic action. The degree of maximum rest is achieved with the opposite actions.
The modulation of the cardioaccelerator and cardioinhibitory centers, mentioned origins of cardiac autonomic innervation, depends on the activity of higher nervous centers located in the brainstem, hypothalamus and cerebral cortex..


Yet No Comments