The DNA packaging is a term that defines the controlled compaction of DNA within the cell. In no cell (and indeed, not even in viruses) is DNA free, loose, and in true solution.
DNA is an extremely long molecule that is also always interacting with a huge variety of different proteins. For the processing, inheritance and control of the expression of the genes it carries, DNA adopts a particular spatial organization. This is achieved by the cell by strictly controlling each step of DNA packaging at different levels of compaction..
Viruses have different packaging strategies for their nucleic acids. One of the favorites is the one of the formation of compact spirals. It could be said that viruses are nucleic acids packaged in their own proteins that cover, protect and mobilize them.
In prokaryotes, DNA is associated with proteins that determine the formation of complex loops in a structure called nucleoid. The maximum level of DNA compaction in a eukaryotic cell, on the other hand, is the mitotic or meiotic chromosome..
The only instance in which a B-DNA is not packaged is a research laboratory that pursues this purpose..
Article index
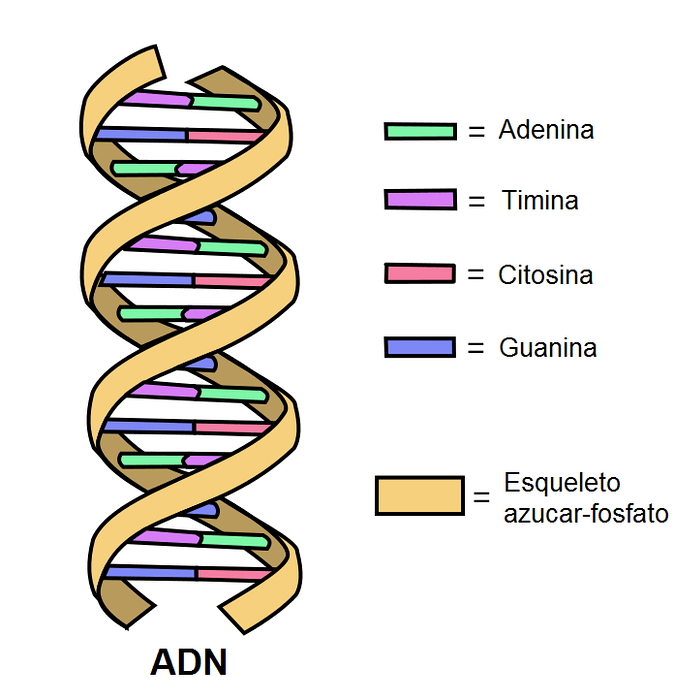
DNA is made up of two antiparallel bands that form a double helix. Each of them has a skeleton of phosphodiester bonds on which sugars linked to nitrogenous bases are attached..
Inside the molecule, the nitrogenous bases of one band form hydrogen bonds (two or three) with the complementary band.
In a molecule like this, most of the important bond angles show free rotation. Nitrogen base-sugar, sugar-phosphate group and phosphodiester bonds are flexible.
This allows DNA, seen as a flexible rod, to show some ability to bend and twist. This flexibility allows DNA to adopt complex local structures and form interaction loops at short, medium and long distances..
This flexibility also explains how 2 meters of DNA can be maintained in each diploid cell of a human being. In a gamete (haploid cell), it would be a meter of DNA.
Although it is not an unbreakable rule, the bacterial chromosome exists as a single supercoiled double-band DNA molecule..
The double helix twists more on itself (more than 10 bp per turn) thus producing some compaction. Local knots are also generated thanks to manipulations that are enzymatically controlled.
In addition, there are sequences in DNA that allow domains to form in large loops. We call the structure resulting from the supercoil and ordered loops a nucleoid..
These undergo dynamic changes thanks to some proteins that provide some structural stability to the compacted chromosome. The degree of compaction in bacteria and archaea is so efficient that there can be more than one chromosome per nucleoid.
The nucleoid compacts the prokaryotic DNA at least 1000 times. The very topological structure of the nucleoid is a fundamental part of the regulation of the genes carried by the chromosome. That is, structure and function constitute the same unit..
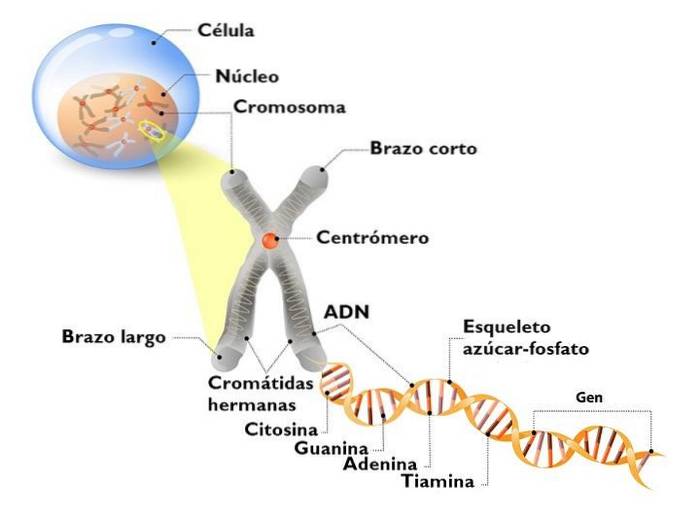
The DNA in the eukaryotic nucleus is not naked. It interacts with many proteins, the most important of which are histones. Histones are small, positively charged proteins that bind to DNA non-specifically.
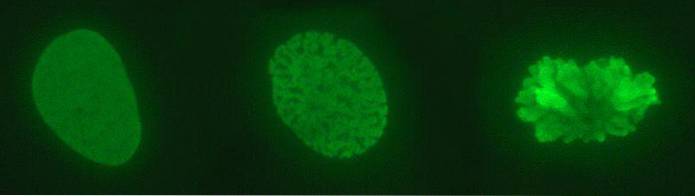
In the nucleus what we observe is a complex DNA: histones, which we call chromatin. The highly condensed chromatin, which is generally not expressed, is heterochromatin. On the contrary, the least compacted (looser), or euchromatin, is chromatin with genes that are expressed.
Chromatin has various levels of compaction. The most elementary is that of the nucleosome; It is followed by the solenoid fiber and the interphase chromatin loops. It is only when a chromosome divides that maximum compaction levels are shown..
The nucleosome is the basic unit of organization of chromatin. Each nucleosome is made up of an octamer of histones that form a kind of drum..
The octamer is made up of two copies of each of the histones H2A, H2B, H3 and H4. Around them, the DNA goes around 1.7 times. This is followed by a fraction of free DNA called a 20 bp linker associated with histone H1, and then another nucleosome. The amount of DNA in a nucleosome and that which binds it to another is about 166 base pairs.
This DNA packaging step compacts the molecule about 7 times. That is, we go from one meter to just over 14 cm of DNA.
This packing is possible because the positive histones cancel the negative charge of the DNA, and the consequent electrostatic self-repulsion. The other reason is that DNA can be folded in such a way that it can flip the octamer of histones..
The fiber of beads in a necklace formed by many successive nucleosomes is further wound into a more compacted structure.
Although we are not clear about what structure it actually adopts, we do know that it reaches a thickness of about 30 nm. This is the so-called 30 nm fiber; Histone H1 is essential for its formation and stability.
The 30 nm fiber is the basic structural unit of heterochromatin. That of lax nucleosomes, that of euchromatin.
The 30 nm fiber, however, is not completely linear. On the contrary, it forms loops of about 300 nm in length, in a serpentine manner, on a little-known protein matrix..
These loops on a protein matrix form a more compact chromatin fiber 250 nm in diameter. Finally, they align as a single helix 700 nm thick, giving rise to one of the sister chromatids of a mitotic chromosome..
Ultimately, the DNA in nuclear chromatin compacts about 10,000 times on the chromosome of the dividing cell. In the interphase nucleus its compaction is also high since it is about 1000 times compared to the "linear" DNA..

In the world of developmental biology, gametogenesis is said to reset the epigenome. That is, it erases the DNA marks that the life of the person who gave rise to the gamete produced or experienced.
These tags include DNA methylation and covalent modifications of histones (Code for histones). But not the entire epigenome is reset. What remains with marks will be responsible for the paternal or maternal genetic imprint.
The implicit reset to gametogenesis is easier to see in sperm. In sperm, DNA is not packed with histones. Therefore, the information associated with its modifications in the producer organism, generally, is not inherited..
In sperm, DNA is packaged through interaction with non-specific DNA-binding proteins called protamines. These proteins form disulfide bonds with each other, thus helping to form overlapping DNA layers that do not repel each other electrostatically..


Yet No Comments