
The DNA translation is the process by which the information contained in the messenger RNAs produced during transcription (the copy of the information in a DNA sequence in the form of RNA) is “translated” into an amino acid sequence by means of protein synthesis.
From a cellular perspective, gene expression is a relatively complex affair that occurs in two steps: transcription and translation..
All genes that are expressed (whether or not they encode peptide sequences, that is, proteins) initially do so by transferring the information contained in their DNA sequence to a messenger RNA (mRNA) molecule through a process called transcription.
Transcription is achieved by special enzymes known as RNA polymerases, which use one of the complementary strands of the gene's DNA as a template for the synthesis of a “pre-mRNA” molecule, which is subsequently processed to form a mature mRNA..
For genes that code for proteins, the information contained in mature mRNAs is “read” and translated into amino acids according to the genetic code, which specifies which codon or nucleotide triplet corresponds to which particular amino acid..
The specification of the amino acid sequence of a protein, therefore, depends on the initial sequence of nitrogenous bases in the DNA that corresponds to the gene and then in the mRNA that carries this information from the nucleus to the cytosol (in eukaryotic cells); process that is also defined as mRNA-guided protein synthesis.
Given that there are 64 possible combinations of the 4 nitrogenous bases that make up DNA and RNA and only 20 amino acids, an amino acid can be encoded by different triplets (codons), which is why the genetic code is said to be "degenerate" (except for the amino acid methionine, which is encoded by a unique AUG codon).
Article index
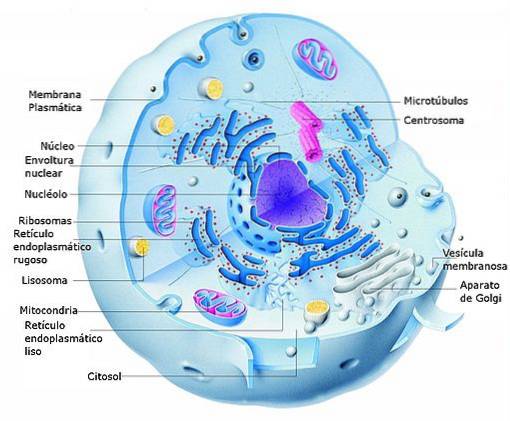
In eukaryotic cells, transcription takes place in the nucleus and translation in the cytosol, so the mRNAs that are formed during the first process also play a role in the transport of information from the nucleus to the cytosol, where the cells are found. biosynthetic machinery (ribosomes).
It is important to mention that the compartmentalization of transcription and translation in eukaryotes is true for the nucleus, but it is not the same for organelles with their own genome such as chloroplasts and mitochondria, which have systems more similar to those of prokaryotic organisms..
Eukaryotic cells also have cytosolic ribosomes attached to the membranes of the endoplasmic reticulum (rough endoplasmic reticulum), in which the translation of proteins that are destined to insert into cell membranes or that require post-translational processing that occur in said compartment occurs..
The mRNAs are modified at their ends as they are transcribed:
- When the 5 'end of the mRNA emerges from the surface of RNA polymerase II during transcription, it is immediately "attacked" by a group of enzymes that synthesize a "hood" composed of 7-methyl guanylate and which is connected to the terminal nucleotide. of the mRNA via a 5 ', 5' triphosphate linkage.
- The 3 'end of the mRNA undergoes a "cleavage" by an endonuclease, which generates a free 3' hydroxyl group to which a "string" or "tail" of adenine residues (from 100 to 250) is attached, which are added one at the same time by an enzyme poly (A) polymerase.
The "hood 5 '" and the "tail poly A ”fulfill functions in the protection of mRNA molecules against degradation and, in addition, they function in the transport of mature transcripts towards the cytosol and in the initiation and termination of translation, respectively..
After transcription, the “primary” mRNAs with their two modified ends, still present in the nucleus, go through a “splicing” process whereby intronic sequences are generally removed and the resulting exons are joined (post-transcriptional processing). , with which mature transcripts are obtained that leave the nucleus and reach the cytosol.
Splicing is carried out by a riboprotein complex called the spliceosome (Anglicism of spliceosome), made up of five small ribonucleoproteins and RNA molecules, which are capable of “recognizing” the regions to be removed from the primary transcript.
In many eukaryotes there is a phenomenon known as "alternative splicing", which means that different types of post-transcriptional modifications can produce different proteins or isoenzymes that differ from each other in some aspects of their sequences..
When mature transcripts leave the nucleus and are transported for translation in the cytosol, they are processed by the translational complex known as the ribosome, which consists of a complex of proteins associated with RNA molecules..
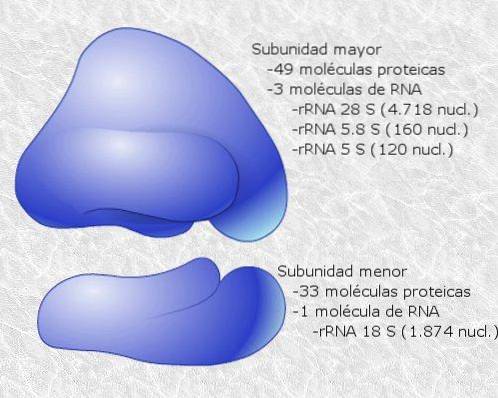
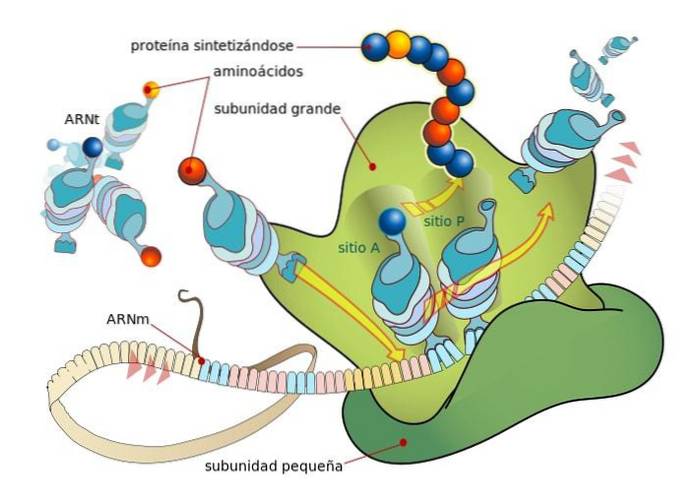
Ribosomes are composed of two subunits, one "large" and the other "small", which are freely dissociated in the cytosol and bind or associate on the mRNA molecule that is translated.
The binding between ribosomes and mRNA depends on specialized RNA molecules that associate with ribosomal proteins (ribosomal RNA or rRNA and transfer RNA or tRNA), each of which has specific functions..
TRNAs are molecular "adapters", since through one of their ends they can "read" each codon or triplet in the mature mRNA (by base complementarity) and through the other they can bind to the amino acid encoded by the "read" codon..
The rRNA molecules, on the other hand, are responsible for accelerating (catalyzing) the binding process of each amino acid in the nascent peptide chain..
A mature eukaryotic mRNA can be "read" by many ribosomes, as many times as the cell indicates. In other words, the same mRNA can give rise to many copies of the same protein..
When a mature mRNA is approached by ribosomal subunits, the riboprotein complex "scans" the sequence of said molecule until it finds a start codon, which is always AUG and involves the introduction of a methionine residue.
The AUG codon defines the reading frame for each gene and, in addition, defines the first amino acid of all proteins translated in nature (this amino acid is often eliminated post-translationally).
Three other codons have been identified as those that induce translation termination: UAA, UAG, and UGA..
Those mutations that involve a change of nitrogenous bases in the triplet that codes for an amino acid and that result in stop codons are known as nonsense mutations, since they cause a premature arrest of the synthesis process, which forms shorter proteins.
Near the 5 'end of mature mRNA molecules there are regions that are not translated (UTR). UnTranslated Region), also called “leader” sequences, which are located between the first nucleotide and the translation start codon (AUG).
These UTR regions that are not translated have specific sites for binding with ribosomes and in humans, for example, are approximately 170 nucleotides in length, among which there are regulatory regions, protein binding sites that function in the regulation of the translation, etc.
Translation, as well as transcription, consists of 3 phases: an initiation phase, an elongation phase, and finally a termination phase..
It consists of the assembly of the translational complex on the mRNA, which merits the union of three proteins known as initiation factors (IF). Initiation Factor) IF1, IF2 and IF3 to the small subunit of the ribosome.
The "pre-initiation" complex formed by the initiation factors and the small ribosomal subunit, in turn, bind with a tRNA that "carries" a methionine residue and this set of molecules binds to the mRNA, near the start codon. AUG.
These events lead to the binding of the mRNA to the large ribosomal subunit, leading to the release of initiation factors. The large ribosome subunit has 3 binding sites for tRNA molecules: the A site (amino acid), the P site (polypeptide), and the E site (exit)..
Site A binds to the anticodon of the aminoacyl-tRNA that is complementary to that of the mRNA being translated; the P site is where the amino acid is transferred from the tRNA to the nascent peptide and the E site is where it is found in “empty” tRNA before being released into the cytosol after the amino acid is delivered.
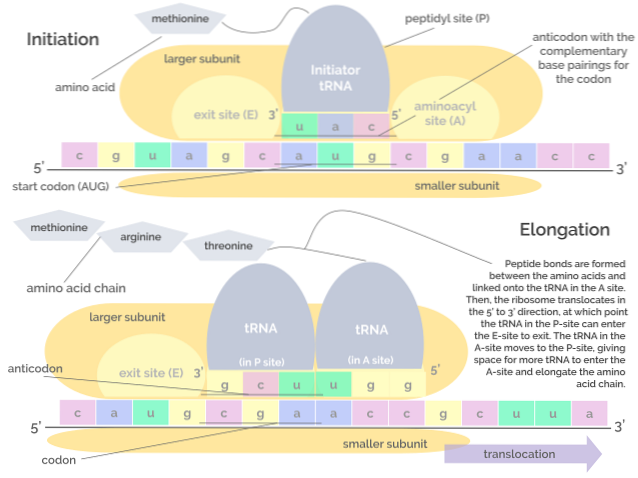
This phase consists of the “movement” of the ribosome along the mRNA molecule and the translation of each codon that is “reading”, which implies the growth or elongation of the polypeptide chain at birth..
This process requires a factor known as elongation factor G and energy in the form of GTP, which is what drives the translocation of elongation factors along the mRNA molecule while it is being translated..
The peptidyl transferase activity of ribosomal RNAs allows the formation of peptide bonds between successive amino acids that are added to the chain.
Translation ends when the ribosome meets any of the termination codons, since tRNAs do not recognize these codons (they do not encode amino acids). Proteins known as release factors also bind, which facilitate the detachment of the mRNA from the ribosome and the dissociation of its subunits..
In prokaryotes, as in eukaryotic cells, the ribosomes responsible for protein synthesis are found in the cytosol (which is also true for the transcriptional machinery), a fact that allows the rapid increase in the cytosolic concentration of a protein when the expression of the genes that encode it increases.
Although not an extremely common process in these organisms, the primary mRNAs produced during transcription can undergo post-transcriptional maturation through "splicing." However, the most common is to observe ribosomes attached to the primary transcript that are translating it at the same time as it is being transcribed from the corresponding DNA sequence..
In view of the above, translation in many prokaryotes begins at the 5 'end, since the 3' end of the mRNA remains attached to the template DNA (and occurs concomitantly with transcription)..
Prokaryotic cells also produce mRNA with untranslated regions known as the "Shine-Dalgarno box" and whose consensus sequence is AGGAGG. As is evident, the UTR regions of bacteria are considerably shorter than those of eukaryotic cells, although they exert similar functions during translation..
In bacteria and other prokaryotic organisms the translation process is quite similar to that in eukaryotic cells. It also consists of three phases: initiation, elongation and termination, which depend on specific prokaryotic factors, different from those used by eukaryotes..
Elongation, for example, depends on elongation factors known as EF-Tu and EF-Ts, rather than the G factor of eukaryotes..



Yet No Comments